


| name | Amanita humboldtii |
| name status | nomen acceptum |
| author | Singer |
| english name | "Humboldt's Ringless Amanita" |
| cap |
The data in this description is almost entirely dependent on the protolog and a study of the type. The cap of Amanita humboldtii is usually free of volval remnants, 35 - 60 mm wide, subumbonate to depressed, with a strongly striate margin; it is dull fuscous brown or ranging from dark orange-brown to fuligineous in the center and orange-brown to medium dark brown in the outer part. [Note: the description seems to mix the present species with A. fuligineodisca Tulloss, Ovrebo & Halling. The reader should assume that the color of A. humboldtii is "dull fuscous brown."] |
| gills |
Gills are free, close, white in side view, and about 5 mm broad, sometimes marginate and then having edge white or concolorous with pileus on the portion of the gills closest to the cap margin. There is no information on the short gills. |
| stem |
The stem is 60 - 180 × 6 - 11 mm, entirely white or white on the lower half and pale orangish tan on the upper half, exannulate, and has a membranous sack-like volva at the base. Singer describes the volval sac as (for example) 20 × 12 mm high. The volval sack may be white, cinnamon, or orange; it is largely free from the stem. |
| spores |
The spores measure (10.2-) 11.0 - 14.0 (-15.5) × (9.0-) 9.8 - 13.0 (-13.3) µm and are inamyloid, globose to subglobose or (infrequently) broadly ellipsoid. Clamps are rare or absent at bases of basidia. |
| discussion |
Amanita humboldtii is known from Andean Colombia, occurring with oak. Apparently, it is not frequently collected. One collection that might be attributable to this species has been sent me from Costa Rica. I have never seen fresh material of this species. Judging from Singer's original description, the species is apparently quite similar macroscopically to A. fuligineodisca Tulloss, Ovrebo & Halling macroscopically, but the two taxa can be distinguished microscopically. Based on the limited number of available collections known, A. humboldtii has larger spores and more layers of cells in its subhymenium. The closest relative of A. humboldtii may be A. pekeoides G. S. Ridl. of New Zealand.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita humboldtii | ||||||||
| author | Singer. 1963. Mycopathol. Mycol. Appl. 20: 241, figs. 8-10. | ||||||||
| name status | nomen acceptum | ||||||||
| english name | "Humboldt's Ringless Amanita" | ||||||||
| etymology | genitive of a Latinized name; hence, "Humboldt's" or "of Humboldt" | ||||||||
| MycoBank nos. | 326098 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| holotypes | BAFC | ||||||||
| revisions | Tulloss et al. (1992) Mem. New York Bot. Gard. 66: 16, figs. 8-10. | ||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text representing a type study by Tulloss. The same field may also contain black text, which will represent a revision of the species by Tulloss. Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following is based upon original research by R. E. Tulloss. | ||||||||
| pileus | 37 - 60 mm diam., “cochin” (MP) (dull fuscous brown) or 6D-F8 in disk and 6C7 or 6E6 toward margin, planoconvex, subumbonate to depressed, slightly viscid when wet; context white, unchanging on exposure, 3.5± mm thick; margin pectinate-sulcate, decurved; universal veil absent. | ||||||||
| lamellae | free, close, white, drying a pale slightly sordid tan or brown (6D-E4), 5± mm broad, with fimbriate edge that may be white or (at least near pileus margin) marginate (concolorous with pileus surface). | ||||||||
| stipe | 63 - 180 × 6 - 11 mm, entirely white or somewhat darker than 5B3 in upper half and otherwise white, firm, narrowing upward, glabrous to fibrillose; context white, solid or becoming hollow especially in lower portion; exannulate; universal veil as free volval sac, “caramel” (MP) (cinnamon to orange) or whitish, membranous, 20± × 12± mm. | ||||||||
| odor/taste | Odorless. Taste not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | 20 - 56 µm thick; slightly gelatinizing subradially arranged interwoven filamentous undifferentiated hyphae with a pale yellowish brown intracellular pigment partially soluble in KOH, 1.2 - 7.8 µm wide; vascular hyphae 1.8 - 11.0 µm wide, branching, relatively common. | ||||||||
| pileus context | filamentous undifferentiated hyphae loosely interwoven to tangled, sometimes in fascicles, 1.2 - 12.8 µm wide, with some intercalary segments slightly inflated to 14.0 µm wide; acrophysalides with walls thin or up to 0.5 µm thick, slenderly clavate to subventricose to ventricose (to 92 × 27 µm) to broadly clavate (to 71 × 42 µm) to ellipsoid to subglobose or pyriform (to 63 × 51 µm); vascular hyphae 1.5 - 10.5 µm wide, branching, common. | ||||||||
| lamella trama | bilateral; wcs = 50 - 55 µm; central stratum very distinct; subhymenial base with angle of divergence 45° - 90°; filamentous, undifferentiated hyphae 2.1 - 11.5 µm wide, branching; inflated cells up to 34 × 20 µm, clavate to ventricose to ellipsoid, diverging at angles cited above; vascular hyphae 2.2 - 6.3 µm wide. | ||||||||
| subhymenium | wst-near = 60 - 70 µm; wst-far = 85 - 105 µm; 4 to 5 inflated cells deep, with basidia arising from uninflated to slightly inflated hyphal segments or (predominantly) from inflated cells of varying size (up to 27 × 25 µm, clavate to broadly clavate to ovoid to ellipsoid to subpyriform to subglobose to globose, arranged in branching chains arising in central stratum). | ||||||||
| basidia | 49 - 78 × 11.5 - 24 µm, thin-walled, clavate, dominantly 4-, occasionally 2-sterigmate; sterigmata to 2.0 µm broad at base; clamps rare. | ||||||||
| universal veil | At stipe base, exterior surface: with very thin layer of filamentous, undifferentiated hyphae 2.2 - 12.6 µm wide, in disorderly loose mat, gelatinized, also occasionally in longitudinally oriented fascicles, dominant; inflated cells up to 39 × 33 µm, gelatinizing; vascular hyphae scarce or absent. At stipe base, interior: filamentous, undifferentiated hyphae 1.4 - 15.8 µm wide, rather densely tangled/interwoven, branching, plentiful, locally dominant, at times in fascicles, somewhat gelatinizing; inflated cells more plentiful than at surfaces, locally dominant, subpyriform to ellipsoid to ovoid to broadly clavate to ellipsoid, thin-walled, to 65 × 54 µm; vascular hyphae 2.8 - 7.0 µm wide. On stipe base, inner surface: as rather thin layer, absent in some specimens or in limited areas of volval limb of other specimens; filamentous, undifferentiated hyphae 1.5 - 8.0 µm wide, branching, longitudinally arranged, partially gelatinizing; inflated cells scattered, collapsing, gelatinizing, up to 52 × 34 µm; vascular hyphae 4.2 - 17.0 µm wide, partially gelatinizing, sublongitudinally arranged to disordered. On pileus: absent. | ||||||||
| stipe context | longitudinally acrophysalidic; filamentous, undifferentiated hyphae 2.0 - 10.5 µm wide; acrophysalides narrowly clavate to narrowly subventricose, up to 290 × 39 µm, with walls thin or up to 0.8 µm thick; vascular hyphae 1.2 - 13.3 µm wide, occasionally tangled in knots. | ||||||||
| basidiospores |
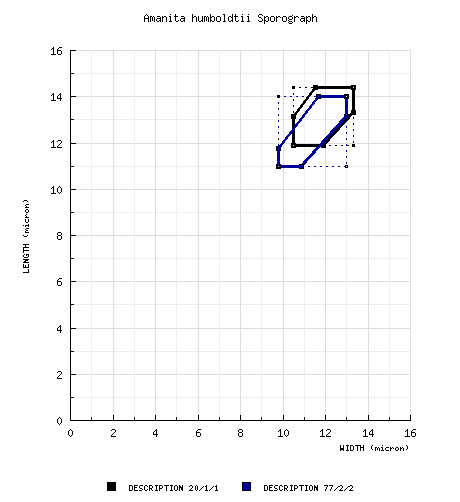
type study, RET: [20/1/1] (10.4-) 11.9 - 14.4 (-15.5) &time; (9.7-) 10.5 - 13.3 μm, (L = 12.8 μm; W = 11.7 μm; Q = 1.0 - 1.25 (-1.28); Q = 1.09),
hyaline, colorless, smooth, thin-walled, inamyloid, globose to subglobose to broadly ellipsoid, often adaxially flattened; apiculus sublateral, cylindric, rather prominent, proportionately broad; contents guttulate; color in deposit not recorded. composite of all RET spore data: [77/2/2] (10.2-) 11.0 - 14.0 (-15.5) × (9.0-) 9.8 - 13.0 ( 13.3) µm, (L = 12.5 - 12.8 µm; L’ = 12.7 µm; W = 11.4 - 11.9 µm; W’ = 11.7 µm; Q = (1.0-) 1.01 - 1.20 (-1.28); Q = 1.08 - 1.09; Q’ = 1.09), inamyloid, hyaline, smooth, globose to subglobose to broadly ellipsoid; contents guttulate to granular, in Melzer’s reagent coagulating and sticking to spore walls; apiculus sublateral, often broad and prominent, cylindrical; color in deposit unknown. | ||||||||
| ecology | Solitary. At 1700 - 2700 m elev. In Quercus humboldtii forests. | ||||||||
| material examined | COLOMBIA: CUNDINAMARCA—Mpio. Pacho - Los Robledales [misspelled in protologue] above Pacho, 26.vii.1960 R. Singer B3527 (BAFC 31.975, holotype); Cerra de Las Mercedes, rd. btwn. Bogotá and La Mesa, s.d. G. Guzmán 4602 (ENCB; COL). NARIÑO—Mpio. Pasto - vereda “La Josefina,” km 17, rd. from Pasto to Chachagüí, 20.xi.1988 A. E. Franco-M. 190 (NY; PSO). | ||||||||
| discussion |
RET has enhanced the macroscopic information in the protologue with data from Singer’s original notes (in F) on the holotype collection as well as with data from the recent collections. The data on anatomy includes that from a complete anatomical review of the holotype. For comparison to A. fuligineodisca see the discussion of that species, directly above. Apparently A. humboldtii is a rather rare species. Amanita humboldtii is a small brownish mushroom with a striate pileus margin, exannulate stipe, and a membranous whitish to cinnamon colored volval sac. In the present state of knowledge, it is distinguished from A. fuligineodisca based on microscopic characters. The specimen Franco-M. 192 (NY; PSO) is probably assignable to this taxon; while it is immature and lacks spores, its anatomy is identical to that of Franco-M. 190 which was collected in the same locality. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita humboldtii |
| bottom links | [ Keys & Checklists ] |
| name | Amanita humboldtii |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.